Historia taxonómica de las hormigas
Fernando Varela Hernández •
Humberto Reyes Prado ••
Introducción
La historia taxonómica de las hormigas ha resultado ser bastante compleja, por lo que las propuestas de relaciones filogenéticas internas se han modificado esencialmente en función del descubrimiento y análisis de nuevos caracteres en cada etapa del desarrollo del conocimiento del grupo. En general, dentro de la historia filogenética de las hormigas se puede reconocer una etapa precladística y una cladística.
En la primera se generaban hipótesis de las relaciones genealógicas de los diferentes grupos de hormigas en función de análisis comparativos de datos empíricos, sobre todo morfológicos y geográficos, pero sin el uso de una metodología rigurosa, por lo que aquellas hipótesis resultaban interpretaciones personales más o menos subjetivas.
En la segunda, la evidencia empírica es interpretada como resultado de un análisis metodológico riguroso. Dentro de esta etapa también se pueden reconocer dos fases, aunque no delimitadas tajantemente pero sí con una tendencia marcada al análisis: primero, de datos morfológicos, y después, de datos moleculares.
A continuación se presenta un breve resumen y recopilación de información desarrollada en diferentes artículos de revistas especializadas sobre la transformación del conocimiento de las relaciones genealógicas (filogenéticas) de las hormigas, desde mediados del siglo pasado hasta la actualidad.
Etapa precladística
Antes de las primeras propuestas filogenéticas hubo un proceso largo de acumulación de conocimiento básico descriptivo de las hormigas (Ward, 2007), lo que sentó las bases para el desarrollo de otros estudios, entre ellos los filogenéticos. Es en 1954 cuando W. L. Brown Jr. hace el primer intento de interpretación de la filogenia de las hormigas, y a partir de entonces se intensifica su estudio en este grupo taxonómico. A partir de 2003 se tiene una revisión de las propuestas filogenéticas de las hormigas (Fernández, 2003), basadas principalmente en caracteres morfológicos, por lo que aquí sólo se consideran algunos aspectos importantes respecto a esas propuestas.
En la primera propuesta formal de filogenia para las hormigas, elaborada en 1954, se reconocen nueve subfamilias pertenecientes a dos grandes grupos: el poneroideo (que incluye a las subfamilias Cerapachyinae, Dorylinae, Leptanillinae, Myrmicinae y Ponerinae) y el mirmecoideo (que incluye a Pseudomyrmecinae, Myrmeciinae, Dolichoderinae y Formicinae). No obstante esta gran aportación inicial sobre las relaciones genealógicas entre subfamilias, las hipótesis derivadas de ella son inciertas, con genealogías dudosas o débilmente respaldadas.
A pesar de esto último, el reconocimiento de estos dos grandes grupos, el poneroideo y el mirmecoideo, aunque por sí mismos no representan ningún nivel taxonómico dentro de la jerarquía lineana, ha sido de gran utilidad como punto de referencia para construir la historia evolutiva de las hormigas.
Otras propuestas interesantes provienen del descubrimiento de hormigas no registradas previamente, tanto fósiles como actuales, así como de su valor predictivo, al ser incluidas dentro de los análisis filogenéticos. En el caso de la hormiga fósil Sphecomyrma freyi (fósil en ámbar), se ha considerado como un ancestro dentro del grupo mirmecoideo (Wilson et al., 1967), pero también se le considera como ejemplar de un grupo hermano de alguno de los dos grandes grupos (Hermann, 1969). No obstante estas diferencias, las hipótesis entre las subfamilias de los dos grandes grupos se mantienen relativamente sin grandes cambios.
Un cambio importante dentro de la filogenia de las hormigas surge con el redescubrimiento de la hormiga australiana Nothomyrmecia (Taylor, 1978), la cual es considerada como una subfamilia aparte (Nothomyrmeciinae) y se le coloca entre Aneuretinae y Sphecomyrminae. Por otro lado, Myrmeciinae y Pseudomyrmecinae pasan a formar parte del complejo poneroide; a las restantes subfamilias se les denomina complejo “formicoide” y se incluye a Sphecomyrma como posible ancestro de Formicidae. De esta manera, en lo subsecuente se abandona el reconocimiento del grupo mirmecoideo y es el formicoideo al que se toma como punto de referencia. Algunas otras propuestas de filogenia enfatizan y reafirman al complejo formicoideo (Dlusskyi y Fodoseeva, 1988), aunque con algunas variaciones internas en las relaciones genealógicas del complejo.
Un aspecto para resaltar es que, hasta aquí, las filogenias de las hormigas incluyen eventos de ancestría-descendencia, más que de relaciones genealógicas. Por ejemplo, se sugería que, dentro del grupo mirmecoideo, Aneuretinae era el grupo ancestral de Dolichoderinae; además, como ya se ha mencionado, se incluía la hormiga fósil como ancestro del grupo mirmecoideo (o de la familia en general), de tal manera que las filogenias resultaban más un aspecto de interpretación que el resultado de un método riguroso.
Etapa cladística (basada principalmente en la morfología)
A finales de los años ochenta se propone por primera vez un análisis cladístico que cambia las posiciones de algunos grupos (Baroni Urbani, 1989). Desde esta perspectiva, el cambio conceptual más importante es el de considerar a los diferentes grupos de estudio como taxones terminales, por lo que sólo se analizan las relaciones de parentesco y no de ancestríadescendencia.
Así, se considera a Sphecomyrma fuera de Formicidae, grupo que pasa a la parte más basal, junto al grupo externo. Por otro lado, aunque se continúan considerando a los complejos poneroideo y formicoideo en general, se proponen cambios internos importantes, por ejemplo, a Pseudomyrmecinae y Myrmicinae se les considera como grupos hermanos, unidos ambos a Myrmeciinae y Ponerinae con ((Dorylinae + Leptanillinae) + Ecitoninae) como su grupo hermano. Finalmente, el resto de las subfamilias se considera como grupo hermano de las anteriores y basales en el cladograma.
Un avance en la filogenia de las hormigas como resultado de este análisis fue el de colocar a los diferentes grupos en posiciones no ambiguas, como se venía haciendo desde los trabajos de Brown (1954). Pese a que se tenía un avance importante en la filogenia de los grandes grupos de hormigas, éste no quedó exento de críticas.
Se cuestionaba básicamente la naturaleza del análisis, debido tanto a la ambigüedad de algunos caracteres que no respaldan la filogenia (además de proponer la separación de Leptanillinae de Dorylinae y la inclusión de Cerapachyinae y Aenictinae como subfamilias) (Bolton, 1990), como por aspectos metodológicos en el análisis, y se concluía que los resultados serían diferentes usando un método más apropiado (Carpenter, 1990, p. 6-8).
Posteriormente, se reanalizan cladísticamente las subfamilias, pero además de las subfamilias reconocidas hasta entonces se integran grupos de hormigas que, por sus características morfológicas, podrían posicionarse en otro clado diferente de aquél donde se les consideraba originalmente (Baroni Urbani et al., 1992). Entre sus resultados más notorios está el reconocimiento de dos grandes grupos: por una parte, el formado por las ponerinas (Apomyrminae, + Ponerinae + Leptanillinae) + Leptanilloidinae, Cerapachyninae, Ecitoninae y (Aenictinae + Aenictogitoninae + Dorylinae); por otra parte, el grupo en el que se encuentran Myrmicinae, Pseudomyrmecinae, Nothomyrmeciinae + Myrmeciinae y (Aneuretinae + Dolichoderinae + Formicinae). Entre las diferencias de clados con respecto a los trabajos anteriores destaca que Ponerinae ya no se encuentra como grupo hermano de Myrmicinae, y Nothomyrmeciinae, que se encontraba como grupo hermano de Dolichoderinae y Formicinae, ahora pasa a formar el grupo hermano de Myrmeciinae (figura 1).
Figura 1
Árbol de consenso estricto

Fuente: Baroni-Urbani et al., 1992.
Como se ha visto, la gran mayoría de las propuestas revisadas hasta entonces están basadas en evidencia morfológica. Con el advenimiento de datos moleculares, principalmente de secuencias de dna, se ha pensado que éstas proporcionarían una base más sólida para la resolución de la filogenia entre las subfamilias. Cabe mencionar que después de algunos trabajos basados en secuencias moleculares, se realiza una filogenia con base en datos morfológicos externos derivados del análisis de microscopía electrónica de barrido (Keller, 2011).
Entre las principales conclusiones se tiene, primeramente, un consenso general respecto a la monofilia de las subfamilias entre datos morfológicos y moleculares, y en segundo lugar, incongruencia entre la topología de ciertas subfamilias, por ejemplo, la posición de Formicinae como grupo hermano de Dolichoderinae de acuerdo con los datos morfológicos. Según los datos moleculares, Formicinae siempre se posiciona como grupo hermano de Myrmicinae. Otro ejemplo sería la posición de Dorylinae, donde esta subfamilia se posiciona como grupo hermano de las mirmicomorfas (Agroecomyrmicinae + Myrmicinae), topología diferente a la de datos moleculares, donde Dorylinae se posiciona como grupo hermano del grupo formicoide.
Etapa cladística (basada principalmente en datos moleculares)
Los primeros análisis moleculares con genes nucleares (28s, 18s, abd-A y Ubx) presentan, por un lado, incongruencia en las relaciones de las subfamilias y, por otro, grupos parafiléticos dentro de los grandes complejos (Ohnishi et al., 2003; Astruc et al., 2004). Por ejemplo, se apoya la hipótesis en cuanto a la posición de Myrmicinae dentro del grupo formicoide y se concuerda en la posición de Ponerinae como grupo hermano del resto de las subfamilias. No obstante, la filogenia no resuelve la posición exacta de las subfamilias implicadas.
Estudios posteriores comenzaron a incluir un mayor número de genes en las filogenias y como resultado se lograron avances importantes. Primeramente, aunque se propusieron cambios en las hipótesis de relaciones entre las subfamilias (por ejemplo, al definir a Leptanillinae como grupo basal de la familia, además de considerar que dentro del grupo formicoide se encuentran Heteroponerinae y Ectatomminae, tradicionalmente caracterizados como pertenecientes a la subfamilia Ponerinae), se comenzó a obtener una estabilidad entre dichas relaciones. En segundo lugar, la incorporación de un mayor número de genes y taxones (Moreau et al., 2006; Brady et al., 2006) en los análisis filogenéticos ha revelado que existe un consenso en cuanto a la monofilia de la mayoría de las subfamilias.
A finales de la década pasada se describe a Martialis heureka sp. nov. (Rabeling et al., 2008) colocándola dentro de un género nuevo y una nueva subfamilia. A través de un análisis filogenético molecular, esta especie queda basal, junto con Leptanillinae, al resto de Formicidae (Moreau y Bell, 2013) (figura 2). Se piensa que ambas subfamilias evolucionaron muy temprano y mantuvieron sus caracteres hasta nuestros días debido a la estabilidad ecológica de sus hábitats.
No obstante, aun cuando la hipótesis basal de estos grupos está bien respaldada estadísticamente, se ha pensado que ciertos factores, como la heterogeneidad en los procesos de substitución de nucleótidos (que generan lo que se conoce como atracción de ramas largas) o la heterogeneidad de la frecuencia de bases nucleotídicas, pueden llevar a interpretaciones filogenéticas erróneas (Ward, 2014).
Figura 2
Esquema de las relaciones filogenéticas entre las subfamilias de hormigas, generado
a través de datos moleculares
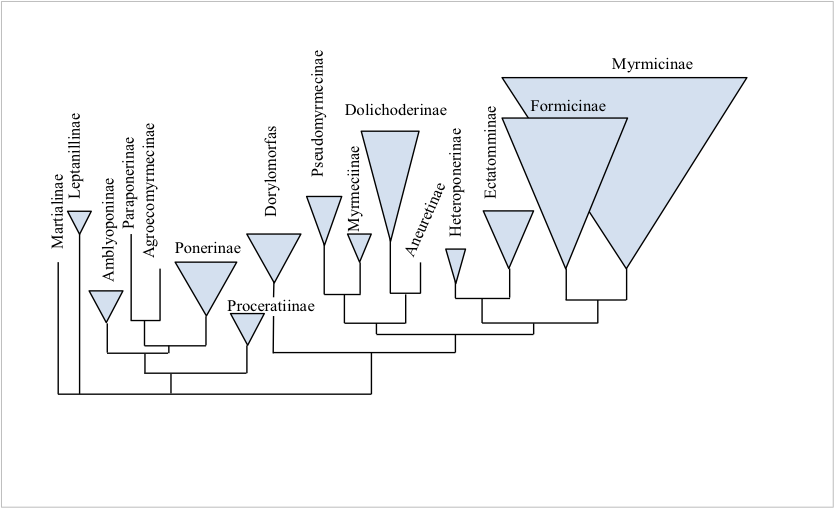
Fuente: Keller, 2011.
En los últimos años se han llevado a cabo análisis filogenéticos menos inclusivos, es decir, estudios cuyos objetivos son los de poner a prueba la monofilia de las subfamilias en concreto, sus fechas de divergencia e historia biogeográfica (Ward et al., 2014; Ward et al., 2016).
Estos trabajos han revelado datos importantes. Por mencionar un ejemplo, dentro la subfamilia Myrmiciane, el género Ankylomyrma pasó a formar parte de la subfamilia Agroecomyrmecinae; además, con el objetivo de una estabilización en la clasificación se sinonimizaron o resucitaron géneros y tribus dentro de la subfamilia. A pesar de esta mejoría en la estabilización de la clasificación interna de esta subfamilia, quedan muchos retos por resolver, como en el caso de la monofilia de los géneros Aphaeonogaster y Monomorium.
Conclusiones
A pesar de que se ha llegado a una relativa estabilidad de las relaciones genealógicas de las hormigas a nivel de subfamilias, aún quedan preguntas qué resolver, ya que, por ejemplo, el registro fósil revela que las hormigas primitivas (Sphecomyrma freyi) presentan un aspecto entre avispa y hormiga. Entre las características evidentes de estas hormigas están la presencia de ojos grandes y supuestamente de hábitos de forrajeo epígeos, de tal manera que en la historia de las hormigas quedarían al menos dos escenarios evolutivos.
Uno es que si las hormigas primitivas eran del tipo de Sphecomyrma, entonces los hábitos hipógeos y la ausencia de ojos de las hormigas basales en las filogenias (esto es, Martialinae y Leptanillinae) habrían sido un carácter derivado (apomorfía), pero si esta hipótesis fuese correcta, entonces todas las subfamilias (aparte de Martialinae y Leptanillinae) tendrían que haber adquirido ojos o hábitos depredadores epígeos de manera secundaria.
El otro escenario es en el que las hormigas primitivas fuesen de hábitos hipógeos, como en Martialinae y Leptanillinae, y que los hábitos epígeos fuesen un carácter derivado. Sin embargo, tendría que incluirse a Sphecomyrma dentro de los grupos actuales de hormigas.
La posición de ciertos clados, como Leptanillinae, genera dudas sobre las posibles secuencias de eventos dentro de Formicidae. En el futuro, tal vez la presencia de un registro fósil más completo, el descubrimiento de nuevas especies o el uso de más información genómica puedan ayudar a un mejor entendimiento de la secuencia de eventos que ha llevado a las hormigas a su éxito ecológico, y de su evolución y diversificación.
• Profesor-investigador, Escuela de Estudios Superiores del Jicarero (eesj), Universidad Autónoma del Estado de Morelos (uaem)
•• Profesor-investigador, Escuela de Estudios Superiores del Jicarero (eesj), Universidad Autónoma del Estado de Morelos (uaem)
Referencias
Baroni Urbani, C. (1989). Phylogeny and behavioral evolution in ants, with a discussion of the role of behavior in evolutionary processes, Ethology, Ecology and Evolution, 1, 137-168.
Baroni Urbani, C., Bolton, B. y Ward, P. S. (1992). The internal phylogeny of ants (Hymenoptera: Formicidae), Systematic Entomology, 17, 301-329.
Bolton, B. (1990). Army ants reassesed: The phylogeny and classification of the doryline section (Hymenoptera:Formicidae). Journal of Natural History, 24, 1339-1364.
Brady, S. G., Schultz, T. R., Fisher, B. y Ward, P. S. (2006). Evaluating alternative hypothesis for the early evolution and diversification of ants. Proceedings of the National Academy of Sciences, 103(48), 18172-18177.
Brown Jr, W. L. (1954). Remarks on the internal phylogeny and subfamily classification of the family Formicidae. Insectes Sociaux, 1, 21-31.
Carpenter, J. M. (1990). On Baroni Urbani’s ant phylogeny. Notes From Underground, 3, 6-8.
Astruc, C., Julien, J.-F., Errard, C. y Lenoir, A. (2004). Phylogeny of ants (Formicidae) based on morphology and sequence data. Molecular Phylogenetics and Evolution, 31, 880-893.
Dlusskyi, G. M. y Fedoseeva, E. B. (1988). Origin and early evolution of the ants. Cretaceous Biocenotic Crisis and Insect Evolution, 70.
Fernández, F. (2003). Sistemática y filogenia de las hormigas: breve repaso a propuestas. Introducción a las hormigas de la región Neotropical. Instituto de Investigaciones de Recursos Biológicos Alexander Von Humboldt.
Hermann, H. R. (1969). The hymenopterous poison apparatus: evolutionary trends in three closely related subfamilies of ants (Hymenoptera: Formicidae). Journal of the Georgia Entomological Society, 4, 123-141.
Keller, R. A. (2011). A phylogenetic analysis of ant morphology (Hymenoptera: Formicidae) with special reference to the poneromorph subfamilies. Bulletin of the American Museum of Natural History, 355, 1-90.
Moreau, C., Bell, C. D., Vila, R., Archibald, B. y Pierce, N. E. (2006). Phylogeny of the ants: diversification in the age of angiosperms. Science, 32, 101-104.
Moreau, C. y Bell, C. D. (2013). Testing the museum versus cradle tropical biological diversity hypothesis: phylogeny, diversification and ancestral biogeographic range evolution of the ants. Evolution, 67, 2240-2257.
Ohnishi, H., Hirotami, I. y Masa-Toshi, Y. (2003). Molecular phylogenetic analysis of ant subfamily relationship inferred from rDNA sequences. Genes and Genetic Systems, 78, 419-425.
Rabeling, C., Brown, J. M. y Verhaagh, M. (2008). Newly discovered sister lineage sheds light on early ant evolution. Proceedings of the National Academy of Sciences, 205, 14913-14917.
Taylor, R. W. (1978). Nothomyrmecia macrops: a living fossil ant rediscovered. Science, 201, 1978, 979-985.
Ward, S. P. (2007). Phylogeny, classification, and species-level taxonomy of ants (Hymenoptera: Formicidae). Zootaxa, 1668, 549-563.
Ward, P. S. (2014). The phylogeny and evolution of ants. The Annual Review of Ecology, Evolution and Systematics, 45, 23-43.
Ward, P. S., Brady, S. G., Fisher, B. L. y Schultz, T. R. (2014). The evolution of myrmicine ants: phylogeny and biogeography of a hyperdiverse ant clade (Hymenoptera: Formicidae). Systematic Entomology, 40, 61-81.
Ward, P. S., Blaimer, B. B. y Fisher, B. L. (2016). A revised phylogenetic classification of the ant subfamily Formicinae (Hymenoptera: Formicidae), with resurrection of the genera Colobopsis and Dinomyrmex. Zootaxa, 4072, 343-357.
Wilson, O. E., Carpenter, M. E., y Brown Jr., W. L. (1967). The first Mesozoic ants. Science, 157, 1038-1040.