Inventio
Vol. 21, núm. 53, 2025
doi: https://doi.org/10.30973/inventio/2025.21.53/4
Taxonomía integrativa: la importancia de usar muchos caracteres para identificar especies
Integrative taxonomy: the importance of using numerous characters to identify species
Leopoldo Andrade Gómez
orcid: 0000-0003-4535-3902, leoango23@gmail.com
Escuela Nacional de Estudios Superiores, Unidad Mérida (enes Mérida), Universidad Nacional Autónoma de México (unam)
Alejandra López Jiménez
orcid: 0000-0002-4032-6094, aleloji@ciencias.unam.mx
Facultad de Ciencias (fc), Universidad Nacional Autónoma de México (unam)
Gerardo Pérez Ponce de León
orcid: 0000-0001-6472-5113, ppdleon@enesmerida.unam.mx
Escuela Nacional de Estudios Superiores, Unidad Mérida (enes Mérida), Universidad Nacional Autónoma de México (unam)
resumen
La práctica taxonómica es inherente a la necesidad del ser humano de nombrar a los objetos y seres vivos de su entorno. Carlos Linneo fundó la disciplina de la taxonomía en un ámbito completamente biológico; además, propuso nombrar las especies bajo un concepto binomial: género y especie. La taxonomía tradicional tuvo que reinventarse bajo nuevos enfoques y tecnologías, incluyendo la incorporación de secuencias de adn. En este sentido, la taxonomía integrativa se enfocó en estudiar la biodiversidad mediante diferentes fuentes de información. En el presente trabajo se repasan los enfoques actuales y los diferentes usos de la taxonomía integrativa, así como las limitantes que ésta presenta.
palabras clave
sistemática, taxonomía, adn, especies, diversidad biológica, morfología
abstract
The practice of taxonomy is inherent to the need of human beings to name the objects and living beings in their environment. Carlos Linneo founded the discipline of taxonomy in a completely biological sphere, addition, proposed naming species under a binomial concept, genus and species. The traditional taxonomy had to be reinvented under new approaches and technologies, including the incorporation of dna sequences. In this sense, the integrative taxonomy focused on studying biodiversity through different sources of information. This paper reviewed the current approaches and the different uses of integrative taxonomy, as well as its limitations.
key words
systematics, taxonomy, dna, species, biological diversity, morphology
Recepción: 03/10/24. Aceptación: 10/02/25. Publicación: 24/06/25.
Introducción
La taxonomía es la rama de la biología que se dedica a la identificación, clasificación y descripción de las especies (Mayr y Ashlock, 1991). La práctica taxonómica es inherente a la necesidad del ser humano de conocer y nombrar a los objetos y seres vivos de su entorno. De esta forma se tiene un panorama más amplio con el cual comunicarnos y entendernos, debido a que hacemos uso de las especies que nos rodean para alimento, vestido, casa, entre otros.
Este deseo de conocer nuestro entorno y describirlo se remonta a la época de los griegos, pasando por la Edad Media y, finalmente, en la edad moderna, a través de la fundación de la disciplina desarrollada por el naturalista sueco Carlos Linneo (1707-1778), quien es considerado el padre de la taxonomía. Linneo describió aproximadamente seis mil especies de plantas y cuatro mil especies de animales, incluyendo el nombre Homo sapiens para la especie humana.
Además de la descripción de muchas especies del planeta, una de las mayores contribuciones de Linneo fue el desarrollo del sistema de nomenclatura binomial. Éste se refiere a que los nombres de las especies estén conformados por dos palabras (binomio), las cuales pertenecen a dos categorías taxonómicas: género y especie (Knapp et al., 2004). Asimismo, se le atribuye el utilizar un lenguaje universal, a través del uso del latín y la implementación de un sistema de organización en el que las categorías taxonómicas se incluyen unas dentro de otras (figura 1).
El principal objetivo de estas clasificaciones era reconocer grupos de organismos que tuvieran características en común. En este sentido, se asumía que las especies que pertenecen a un género en particular provienen de un ancestro común; por lo tanto, compartirán ciertas particularidades. Por ejemplo, se reconocen entre diez y doce especies pertenecientes al género Homo, aunque casi todas están extintas. Algunas de ellas son, por ejemplo, Homo sapiens, H. erectus, H. habilis y H. neanderthalensis, entre otras. El género Homo indica que todas estas especies provienen de un ancestro común.
Si hacemos una analogía, el género correspondería al apellido y la especie al nombre. En este ejemplo, el apellido une a los integrantes de una familia y el nombre distingue a cada uno de ellos (Argue et al., 2017; Wood, 1992). Se puede generalizar que una especie es considerada como un conjunto de organismos que provienen de un ancestro en común y que evolucionan de manera separada de otros grupos de organismos. No obstante, el concepto de especie es un tema que ha generado debate durante muchos años y probablemente nunca se llegue a un acuerdo dentro de la comunidad científica; basta con mencionar que existen más de cien definiciones de especie (Zachos, 2016).
En cuanto a la pregunta acerca de por qué denominar a las especies con nombres curiosos y en latín, la respuesta es que se nombran así para que cualquier persona pueda identificar y saber de qué organismo se trata. Los nombres de las especies son asignados por el taxónomo que las describe por primera vez; en ocasiones se utiliza alguna característica en particular que posee la especie. Por ejemplo, puede ser algún rasgo sobre su morfología, o algún aspecto ecológico, o quizá se haga referencia al lugar en el que se encontró la especie, entre otras. Es decir, el nombre de la especie nos permite tener una idea de cómo podría ser ésta o dónde podría estar ubicada.
Un ejemplo claro podemos encontrarlo en el elefante africano de la sabana, cuyo nombre científico es Loxodonta africana. En primer lugar, el epíteto específico, africana, nos indica que es un organismo que se encuentra distribuido en África. Retomando el ejemplo del Homo sapiens, el género Homo se refiere a un homínido, y sapiens, a la cualidad de inteligente, es decir, es un homínido inteligente. Pero ¿qué tal la especie Darthvaderum greensladeae? Este ácaro es famoso debido a que la parte anterior de su cuerpo se asemeja a la máscara de Darth Vader, personaje de las películas de la serie La guerra de las galaxias; a su vez, greensladeae hace referencia a que se trata de una especie que se describió en honor a Penelope Greenslade, investigadora experta en artrópodos.
Otro ejemplo es el del hongo Spongiforma squarepantsii que, como su nombre lo indica, se describió así en honor al personaje de la serie de televisión animada Bob Esponja. De acuerdo con los autores, se le nombró de esta manera debido a la forma del cuerpo del hongo, así como por la presencia de filamentos que se asemejan a pequeñas esponjas. Por lo tanto, de manera general, los nombres científicos de las especies nos indican características particulares de los organismos observados (Cuvier, 1827; Desjardin et al., 2011; Hunt, 1996).
Figura 1
Principales categorías taxonómicas
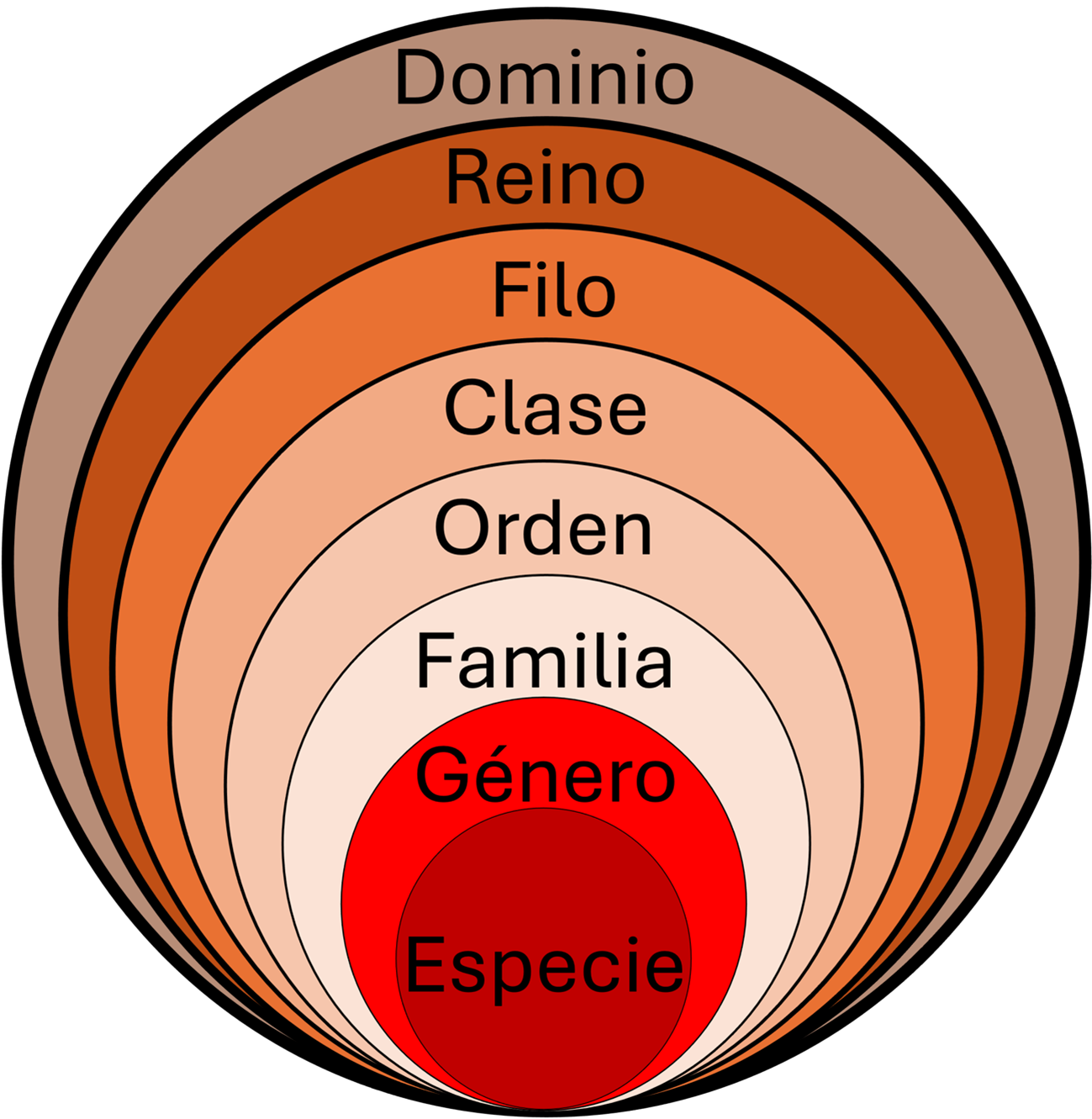
Fuente: elaboración propia.
Figura 2
Esquema filogenético con cuatro especies hipotéticas
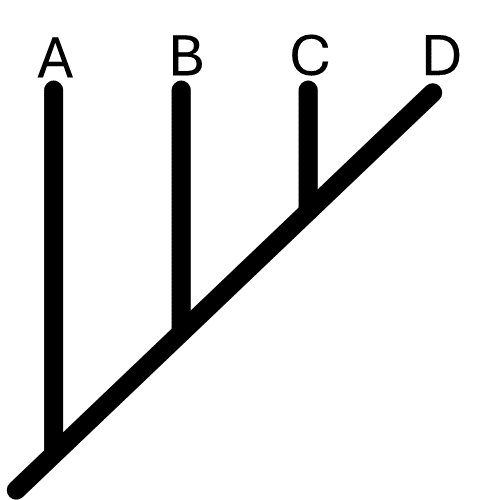
Fuente: elaboración propia.
Taxonomía integrativa
Hace algunos años se reconoció que la biología debía tener un enfoque multidisciplinario (Wake, 2003). Por lo general, ésta se consideraba tan sólo como una disciplina descriptiva y no eran claros los alcances de la información que se producía de la práctica taxonómica. En los últimos veinte años, la taxonomía ha tenido que reinventarse, por lo que, en 2005, un investigador estadounidense especialista en el estudio taxonómico de moluscos, Benoît Dayrat, publicó un trabajo donde definió la taxonomía integrativa de manera puntual como una aproximación al estudio de la diversidad en la que se consideran diferentes fuentes de información, lo que le confiere precisamente un carácter multidisciplinario.
Dayrat (2005) definió la taxonomía integrativa, entonces, como “una disciplina que ayuda a delimitar las unidades de la diversidad de la vida desde perspectivas múltiples y complementarias”. Por lo tanto, esta disciplina tiene como objetivo delimitar, y a partir de ello nombrar, a las especies utilizando diferentes fuentes de evidencia, así como proveer la información necesaria para poder clasificarlas. Esta forma de aproximar la descripción de la biodiversidad surgió como un complemento a la práctica de la taxonomía tradicional, aquella que se basa sólo en los caracteres morfológicos de los organismos.
La morfología demostró ser una fuente de evidencia que, si bien ha sido la base del reconocimiento de las especies, tiene la desventaja de que, en ocasiones, la misma variación entre los individuos se convierte en un reto para explicar cuándo estamos tratando con dos entidades diferentes. Por ejemplo, en las cícadas, que son plantas gimnospermas, es decir, que tienen semillas desnudas, como los pinos, se ha observado que la especie Zamia paucijuga presenta gran variación en el tamaño de sus foliolos (hojas modificadas). Si sólo nos basamos en ese carácter, podríamos concluir erróneamente que cada forma distinta del foliolo de esta especie correspondería a una especie diferente (Nicolalde-Morejón et al., 2019). Por lo anterior, es de suma importancia utilizar más de una evidencia al momento de delimitar y describir a las especies, es decir, tener múltiples fuentes de información.
En este sentido, la taxonomía integrativa utiliza diferentes fuentes de evidencia para generar hipótesis robustas y, de esa forma, delimitar adecuadamente a las especies. En años recientes, diversos estudios enfocados en la taxonomía integrativa han recurrido al uso de secuencias de adn, también conocidas como marcadores moleculares. Estos marcadores han permitido contrastar, robustecer y complementar la taxonomía tradicional. Los marcadores moleculares son secuencias del adn nuclear o mitocondrial que se utilizan normalmente bajo un enfoque filogenético, esto es, las secuencias se comparan con otras parecidas y se generan árboles filogenéticos que nos permiten inferir la historia evolutiva de las especies, a partir de lo cual un taxónomo puede construir un sistema de clasificación (figura 2) (Avise, 1994).
Adicionalmente, con las secuencias de adn se calcula la divergencia genética con respecto a las otras especies analizadas, es decir, se realiza una estimación de las diferencias genéticas entre los individuos, que es una forma de establecer límites entre ellos para poder decidir si son especies diferentes; por ello, esta fuente de caracteres resulta ser complementaria de la morfología. En algunas ocasiones los datos moleculares informan sobre la existencia de una especie que no ha sido descrita con anterioridad y que no puede ser diferenciada de las demás morfológicamente, pero que está separada genéticamente: éstas también son conocidas como especies crípticas. Sin embargo, esto debe tomarse con precaución, porque los datos moleculares pueden fallar en la separación entre especies cuando el taxónomo ha encontrado suficiente evidencia morfológica para considerar que son dos especies distintas. Por lo tanto, la taxonomía tradicional basada en los caracteres morfológicos de los organismos se complementa con el enfoque de la biología molecular.
Además de los caracteres morfológicos y moleculares como fuentes de evidencia para sustentar las hipótesis de especies, la taxonomía integrativa abarca otras disciplinas para su validación. Los aspectos ecológicos también son de suma importancia para reconocer diferencias entre especies. Por ejemplo, se ha observado que los parásitos tienen una estrecha relación con sus hospederos, que es referida como especificidad hospedatoria; esta asociación ecológica per se es otra evidencia para diferenciar a una especie de otra (Pérez-Ponce de León et al., 2016).
Asimismo, la distribución geográfica de las especies es otro tipo de evidencia que nos permite discernir entre especies. La biogeografía es la disciplina encargada de estudiar la distribución de las especies, pues éstas se encuentran en una determinada región geográfica, ya sea por eventos históricos o ecológicos. Otras fuentes de evidencia implementadas en la taxonomía integrativa en diferentes grupos de organismos abarcan desde la bioacústica en aves y anfibios y los modelos de distribución en reptiles, hasta los compuestos químicos generados por diferentes insectos, ciclos de vida en moluscos, entre otros (Alström et al., 2015; Rakotoarison et al., 2017). Sin embargo, la gran mayoría de los estudios se han enfocado en secuencias de adn, la mayoría bajo un contexto filogenético, como se vio anteriormente.
No obstante, aunque la taxonomía integrativa utiliza la mayor cantidad de información disponible sobre una especie para delimitarla, es importante hacer notar sus alcances. Por ejemplo, en ocasiones las distintas fuentes de evidencia se contradicen entre sí. Esto quiere decir que algún carácter no concuerda con los demás analizados. Por lo anterior, se han realizado dos diferentes métodos de integración dentro de la taxonomía: el primero es por acumulación y el segundo por congruencia. El enfoque de integración por acumulación separa especies con algún grado de divergencia en uno o más caracteres taxonómicos, por ejemplo, adn o morfología. Es decir, asume que la diferencia en cualquier característica entre los individuos constituye un carácter taxonómico que puede aportar evidencia suficiente para separar especies (De Queiroz, 2007). De este modo, los taxónomos pueden seleccionar el conjunto de caracteres taxonómicos más apropiados para cada grupo de organismos. Por otro lado, el enfoque de integración por congruencia separa especies combinando dos o más caracteres taxonómicos, por ejemplo, caracteres moleculares, como el adn, más la morfología, con el objetivo de obtener concordancia entre los diferentes conjuntos de información.
DeSalle et al. (2005) elaboraron un esquema donde se ilustra que la congruencia entre dos caracteres taxonómicos o más es un factor importante para determinar la validez de una especie (figura 3). La principal ventaja del enfoque de integración por congruencia es la incorporación de diferentes caracteres taxonómicos, lo que proporciona una mayor confianza en la validez de las especies. No obstante, ambos métodos tienen limitantes al momento de estimar el número de especies. El primer método puede sobreestimar el número de especies al identificar especies distintas cuando probablemente exista variación en algún carácter. Por ejemplo, las poblaciones coespecíficas pueden ser muy distintas desde el punto de vista morfológico, pero sería erróneo considerarlas especies distintas. Por el contrario, el segundo método es un enfoque muy estricto que podría subestimar el número de especies al no poder detectar especies crípticas —morfológicamente similares pero genéticamente diferentes— o recientemente separadas, debido a que el proceso de especiación no siempre va acompañado de un cambio de carácter en todos los niveles (Padial et al., 2010).
Figura 3
Círculo taxonómico
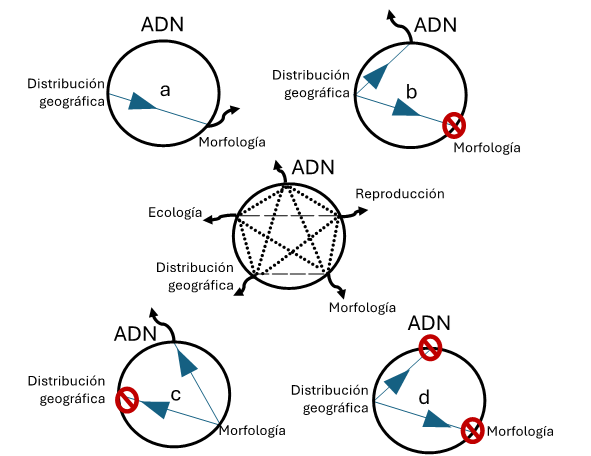
Las líneas discontinuas en la parte interior del círculo indican las vías que pueden seguirse para corroborar a las especies. La única manera de delimitar un nuevo taxón es salirse del círculo. a) Morfología con distribución geográfica. b) Especies crípticas: la distribución geográfica se comprueba con la morfología. La morfología no corrobora la hipótesis geográfica, por lo que el taxónomo no puede salirse del círculo. Manteniendo la hipótesis geográfica, el taxónomo examina secuencias de adn y se produce la corroboración. c) Especies simpátricas: no hay patrón geográfico en la distribución de los diferentes tipos morfológicos, por lo que el taxónomo utiliza secuencias de adn para probar la hipótesis morfológica; así es como corrobora la hipótesis morfológica, con lo que el taxónomo sale del círculo. d) No hay detección de un nuevo taxón: la hipótesis geográfica se comprueba con morfología. La información morfológica falla, la hipótesis geográfica y la hipótesis morfológica se prueban con información de secuencias de adn. La información de adn no permite rechazar la hipótesis de la ausencia de nuevos taxones; por lo tanto, el taxónomo no puede salir del círculo y la conclusión es que sólo existe un taxón.
Fuente: modificado de DeSalle et al. (2005).
Figura 4
Delimitación del concepto de especie a través de diferentes caracteres
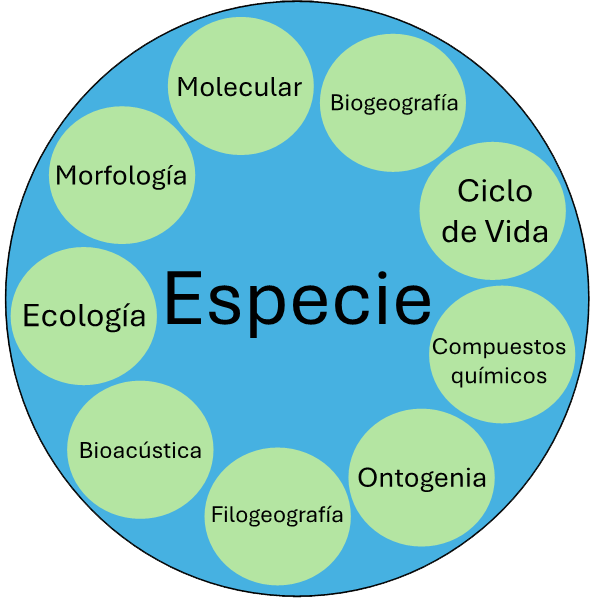
Fuente: elaboración propia.
Conclusiones
La taxonomía integrativa proporciona un enfoque pluralista que agrega la mayor cantidad de fuentes de evidencia para delimitar a las especies, lo que tiene consecuencias para la descripción de la biodiversidad y para la construcción de esquema de clasificación como parte del árbol de la vida en el planeta. Para ello, utiliza todos los caracteres posibles, como los morfológicos, moleculares, ecológicos, biogeográficos, compuestos químicos, bioacústica, modelos de distribución, filogeografía, ontogenia, entre otros (figura 4). Es decir, toda la información que la especie pueda proporcionar es utilizada de manera complementaria, pero al mismo tiempo estos caracteres se contrastan entre ellos para, de este modo, circunscribir cada especie. La taxonomía integrativa aborda las limitantes de utilizar un solo carácter a través de un enfoque más completo y preciso, es decir, de manera holística. Por lo tanto, promueve una comprensión más profunda de la diversidad biológica y permite entender las relaciones evolutivas entre las especies. Por lo anterior, alentamos al lector a continuar estudiando la biodiversidad desde un enfoque integral.
Referencias
Alström, P., Xia, C., Rasmussen, P. C., Olsson, U., Dai, B., Zhao, J., Leader, P. J., Carey, G. J., Dong, L., Cai, T., Holt, P. I., Le Manh, H., Song, G., Liu, Y., Zhang, Y. y Lei, F. (2015). Integrative taxonomy of the russet bush warbler Locustella mandelli complex reveals a new species from central China. Avian Research, 6, 9, 1-33. https://doi.org/10.1186/s40657-015-0016-z
Argue, D., Groves, C. P., Lee, M. S. Y. y Jungers, W. L. (2017). The affinities of Homo floresiensis based on phylogenetic analyses of cranial, dental, and postcranial characters. Journal of Human Evolution, 107, 107-133. https://doi.org/10.1016/j.jhevol.2017.02.006
Avise, J. C. (1993). Kinship and intraspecific phylogeny. En Molecular markers, natural history and evolution (pp. 190-251). Springer. https://books.google.com.mx/books?id=aWSbZf-dRbEC&printsec=frontcover&source=gbs_ge_summary_r&cad=0#v=onepage&q&f=false
Cuvier, F. (1827). L’elephant d’Mrique. En Histoire naturelle des mammifères (pp. 51-52), París, 3(5), 51-52.
Dayrat, B. (2005). Towards integrative taxonomy. Biological Journal of the Linnean Society, 85(3), 407-417. https://doi.org/10.1111/j.1095-8312.2005.00503.x
DeSalle, R., Egan, M. G. y Siddal, M. (2005). The unholy trinity: taxonomy, species delimitation and dna barcoding. Philosophical Transactions of the Royal Society B, 360, 1905-1916. https://doi.org/10.1098/rstb.2005.1722
De Queiroz, K. (2007). Species concepts and species delimitation. Systematic Biology, 56(6), 879-886. https://doi.org/10.1080/10635150701701083
Desjardin, D. E., Peay, K. G. y Bruns, T. D. (2011). Spongiforma squarepantsii, a new species of gasteroid bolete from Borneo. Mycologia, 103, 1119-1123. https://doi.org/10.3852/10-433
Hunt, G. S. (1996). Description of predominantly arboreal platermaeoid mites from eastern Australia (Acarina: Cryptostigmata: Plateremaeoidea). Records of the Australian Museum, 48, 303-324.
Knapp, S., Lamas, G., Lughadha, E. N. y Novarino, G. (2004). Stability or stasis in the names of organisms: the evolving codes of nomenclature. Philosophical Transactions of the Royal Society of London, 359, 611-622. http://doi.org/10.1098/rstb.2003.1445
Mayr, E. y Ashlock, P. D. (1991). Principles of systematic zoology. McGraw-Hill.
Nicolalde-Morejón, F., Martínez-Domínguez, L., Stevenson D. W. y Vergara-Silva, F. (2019). Disentangling the identity of Zamia from Mexican Pacific seaboard, with a description of a new species. Nordic Journal of Botany, 37(9), 1-9. https://doi.org/10.1111/njb.02430
Padial, J. M., Miralles, A., De la Riva, I. y Vences, M. (2010). The integrative future of taxonomy. Frontiers in Zoology, 7, 2-14. https://doi.org/10.1186/1742-9994-7-16
Pérez-Ponce de León, G., García-Varela, M., Pinacho-Pinacho, C. D., Sereno-Uribe A. L. y Poulin, R. (2016). Species delimitation in trematodes using dna sequences: Middle-American Clinostomum as a case study. Parasitology, 143(13), 1773-1789. https://doi.org/10.1017/S0031182016001517
Rakotoarison, A., Scherz, M. D., Glaw, F., Köhler, J., Andreone, F., Franzen, M., Glos, J., Hawlitschek, O., Jono, T., Mori, A., Ndriantsoa, S. H., Raminosoa, N. R., Riemann, J. C., Rödel, M.-O., Rosa, G. M., Vieites, D. R., Crottini, A. y Vences, M. (2017). Describing the smaller majority: integrative taxonomy reveals twenty-six new species of tiny microhylid frogs (genus Stumpffia) from Madagascar. Vertebrate Zoology, 67(3), 271-398. https://doi.org/10.3897/vz.67.e31595
Wake, M. H. (2003). What is “integrative biology”? Integrative and Comparative Biology, 43(2), 239-241. https://doi.org/10.1093/icb/43.2.239
Wood, B. (1992). Origin and evolution of the genus Homo. Nature, 355, 783-790. https://doi.org/10.1038/355783a0
Zachos, F. E. (2016). Species concepts in biology. Historical development, theoretical foundations and practical relevance. Springer.